Previous Lecture | Syllabus | Next Lecture
![]()

Genetic exchange between viruses can only occur when two viruses simultaneously infect the same cell. Then genetic exchange can occur by:
Recombination
This is the breakage and reunion of homologous regions in the nucleic acid molecules from two viruses.
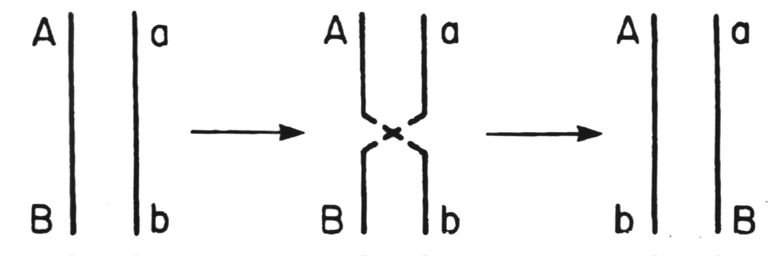
This process occurs within almost all groups of DNA-containing viruses and within the Retroviridae at the DNA provirus stage. The genomes of the Picornaviridae are the only RNAs known to undergo crossovers among animal viruses.
Reassortment
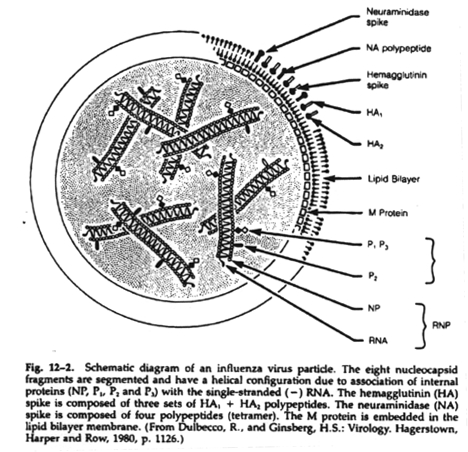
This is an exchange of nucleic acid segments between viruses with segmented genomes. This includes only certain RNA-containing viruses (Orthomyxoviridae, Arenaviridae, Reoviridae and Bunyaviridae). Reassortment is the mechanism of "genetic shift" whereby influenza viruses rapidly acquire new hemagglutinin and neuraminidase antigens. This is the initiating factor in many epidemics.
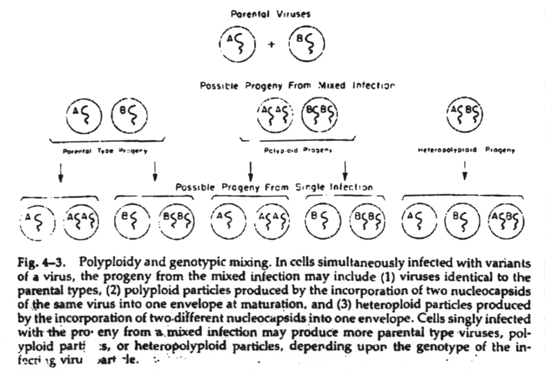
Polyploidy and heteropolyploidy (genotypic mixing)
This is the incorporation of more than one complete genome into the same virus capsid. If the genomes are from the same virus species, the phenomenon is polyploidy; if the genomes are from different virus species, the phenomenon is heteropolyploidy or genotypic mixing. There is no recombination between the genomes and cells singly infected with the genotypically mixed viruses will yield progeny identical to both original parents as well as more genotypically mixed viruses.
Part of the genetic material of a donor cell can be transferred to a recipient cell. After the transfer, recombination between the donor and recipient DNA may occur followed by succeeding nuclear and cell division.
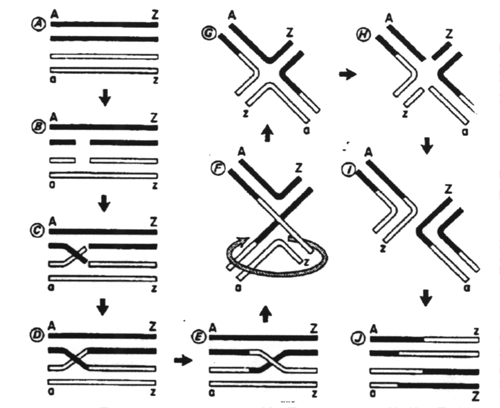
The model for recombination is as follows:
Breakage of two strands occurs.
The broken segments are then reciprocally rejoined.
The crossover point is laterally displaced.
The other strands are broken and partially digested.
Localized repair and reciprocal joining occurs.
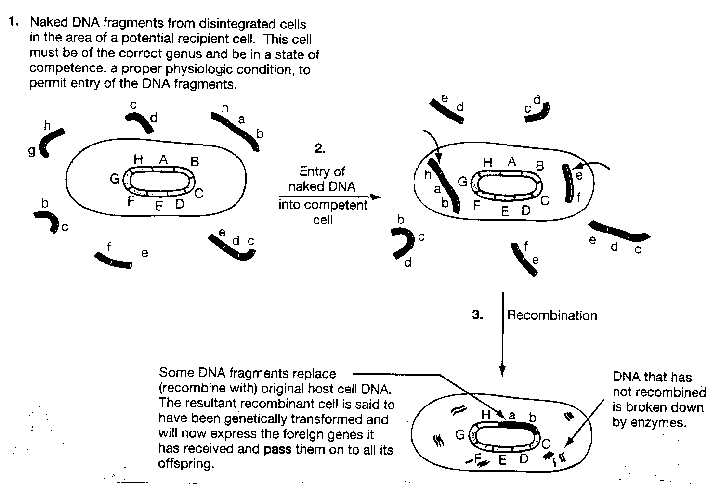
Transformation
This is the uptake of extracellular DNA by bacteria. This process occurs only in bacteria that can take-up high molecular weight DNA from the medium. Most bacterial cells need to be at a particular stage in their growth cycle or under a particular growth regimen in order to be transformed. Also, certain factors such as DNA binding proteins of the cell envelope and poly-b-hydroxybutyrate are necessary for transformation.
Transformation is a three step process:
2. The bound DNA is taken up through the cell membrane.
3.
The donor DNA fragment is then integrated into the host
chromosome or replicates autonomously as a plasmid.
The Gram-positive species that are able to take up exogenous DNA include Streptococcus pneumoniae, Staphylococcus aureus and Bacillus subtilis. Gram-positive strains take up both homologous and heterologous DNA.
Plasmid DNA may also be taken up by competent cells. However, these molecules must be linearized before cell entry. Plasmids may be restored by recombination between overlapping plasmid molecules or recombination with the recipient's chromosome.
Transformation in Gram-negative bacteria
Gram-negative bacteria that can be transformed by exogenous DNA include Neisseria meningitis, Neisseria gonorrhea, Haemophilus influenzae, and Escherichia coli. Homologous DNA is taken up at a much higher rate than heterologous DNA. Transformasomes, membrane bound organelles, sequester DNA and transfer it into the cell's interior. Degradation of one DNA strand follows and only a single strand participates in recombination. Uptake specificity may depend upon specific sites in the donor DNA. Example - Haemophilus influenzae contains 11 base pair sequences that are essential for recognition and DNA uptake.
Transduction
This genetic transfer occurs in both Gram-positive and Gram-negative bacteria when a fragment of DNA is carried to the recipient cell by virus (bacteriophage) produced by a donor cell. Transduction is observed with temperate bacteriophages (those that can form prophages). A prophage is a bacterial virus that has integrated its DNA into the DNA of a bacterial cell. This process of integration of viral DNA into bacterial DNA is lysogenization. Lysogenization that results in a change in the phenotype of the host cell is called lysogenic conversion. Restricted (specialized) transduction - this occurs when the transducing phage only carries segments of DNA that are immediately adjacent to the site of prophage attachment. After phage is introduced into the cell, its DNA becomes integrated into the bacterial chromosome. When the phage becomes induced, the phage DNA is excised from the bacterial chromosome, the phage replicates, and the host cell lyses, releasing mature phage particles. Occasionally, the excision of the phage DNA is imprecise and the resulting excised piece of phage contains some of the host bacterial genome. If the host DNA replaces essential phage genes, the resulting phage will be a defective phage which cannot mature and replicate unless in the presence of a normal lambda phage. Defective phage are also called transducing particles.
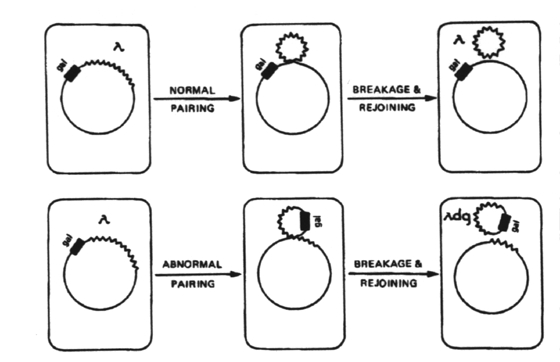
When transducing particles infect donor cells, the donor DNA integrates into the bacterial chromosome in the normal fashion. These transduced genes are expressed in the recipient cell even though they are contained within the prophage DNA.
Specialized transduction occurs when a gene or a set of genes has a high frequency of transduction relative to the majority of genes on the bacterial chromosome.
Plasmids are small autonomously replicating circular pieces of DNA. These may carry genes for resistance to drugs or virulence factors. Many of these plasmids also mediate gene transfer, resulting in bacterial strains with unique drug resistance patters or novel virulence factors. They contain double-stranded circular DNA.
Plasmid transfer in Gram-negative bacteria occurs only between strains of the same species or closely-related species.
Many plasmids of Gram-negative bacteria are conjugative. These plasmids carry genes that mediate their own transfer. These genes code for the production of the sex pilus and enzymes necessary for conjugation.
Conjugation begins with the extrusion of a sex pilus; the tip of the sex pilus adheres to the outer membrane of Gram-negative cell walls. Following pilus adherence, the two cells become bound together at a point of direct envelope-to-envelope contact.
After pair formation, the plasmid will undergo a specific type of replication called "transfer replication." One parental strand of DNA is broken and passes into the recipient while the other remains circularized and in the donor. The complementary strands are synthesized in both donor and recipient cells. Daughter DNA plasmids are recircularized with ligase immediately after transfer replication is complete.
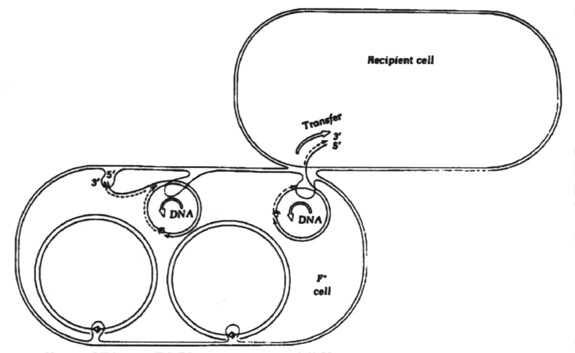
During conjugation, no cytoplasm or cell material except DNA passes from donor to recipient. After conjugation, the cells break apart and two plasmid containing cells result.
Self-transfer in Gram-positive bacteria
Plasmids may integrate into the bacterial chromosome depending upon the extent of DNA homology between the two. After integration, both plasmid and chromosome will replicate as a single unit. A plasmid that is capable of integrating into the chromosome is called an episome.
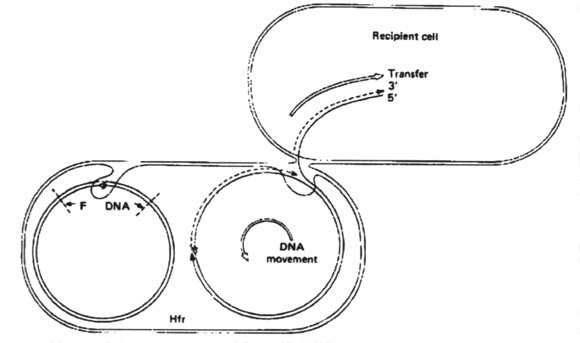
Some conjugative plasmids are able to integrate into the host chromosome. After integration, both chromosome and plasmid can be conjugally transferred to a recipient cell. Plasmids that are able to mobilize chromosomal transfer are called sex factors or F (fertility) factors. Cells that contain the sex factor F are designated F+ and those that do not contain the factor are F-. If the F-plasmid is integrated into the chromosome it is called an Hfr cell.
When Hfr cells are added to an excess of F- cells, all Hfr cells will attach to the susceptible F- cells and replicative transfer (conjugation) will occur. The chromosome and the F factor will be transferred to the recipient.
Cell
Properties Carried by Plasmids
Drug resistance
Gram-negative bacteria carry plasmids that give resistance to antibiotics such as neomycin, kanamycin, streptomycin, chloramphenicol, tetracyclines, penicillins and sulfonamides. Gram-positive Staphylococcus aureus carries plasmids that contain genes for resistance to penicillin, heavy metals (Hg or Co) and erythromycin. Most antibiotic resistance in such strains is mediated by enzymes that inactivate the drug by acetylation or phosphorylation. Chromosomally encoded antibiotic resistance genes often act by altering the binding site for the antibiotic.
Gram-negative plasmids that contain antibiotic resistance genes are called R factors. The R factors are composed of the resistance transfer factor (RTF) segment of the plasmid that contains the genes responsible for intercellular transfer and R determinant that carries the resistance genes.
Virulence
(1)
Toxins
- Enterotoxins (Escherichia coli, Vibrio cholerae), exfoliative
toxin (Staphylococcus aureus), dermotoxin of
Bacillus anthracis, the neurotoxin of Clostridium tetani,
and the pesticide toxin of Bacillus thuringiensis.
(2)
Adhesins
- such as produced by the plasmids of Yersinia enterocolitica,
Shigella
flexneri, Escherichia coli strains that
produce dysentery, and Yersinia pestis.
(3)
Growth
factors - Other plasmid borne virulence factors act to directly aid
the bacteria in competing with mammalian
host cells for growth. For examples, the plasmid Col V of Escherichia
coli contains genes for iron sequestering
compounds. The acquisition of iron is essential for the survival of Escherichia
coli in mammalian infections.
Production of antimicrobial agents
Bacteriocins are a special class of antimicrobics that are active only against other strains of the same species that produced them. A specific example is the colicins which are produced by Escherichia coli cells that harbor small nonconjugative plasmids. These plasmids are called colfactors and contain genes for the colicin it produces, as well as a gene for a protein that protects the donor cell from the colicin. Some colicins act by forming ion-permeable channels in the membrane of sensitive cells. These act to collapse the membrane potential of the colicin sensitive cell.
Metabolic activities
Examples of these genes are those that allow bacteria to utilize unique or unusual materials for carbon or energy sources. Many of the genes for these metabolic pathways are on transmissible plasmids.
|
|
|
| Pseudomonas spp | Degradation of camphor, toluene, octane, salicylic acid |
| Bacillus stearothemophilus | a-Amylase |
| Alcaligenes eutrophus | Utilization of H2 as oxidizable energy source |
| Escherichia coli | Sucrose uptake and metabolism, citrate uptake |
| Klebsiella spp | Nitrogen fixation |
| Streptococcus (group N) | Lactose utilization, galactose phosphotransferase system, citrate metabolism |
| Rhodospirillum rubrum | Synthesis of photosynthetic pigment |
| Flavobacterium spp | Nylon degradation |
Surface antigens
Genetic Exchange Between Fungi
Classes
of fungi based on sexual spore formation
Classes
of fungi based on sexual spore formation
Phycomycetes (Mucor sp. and Rhizopus sp.)
Ascomycetes (Aspergillus sp. and Penicillium sp.)
Basideomycetes (mushrooms)
In the phycomyces, sexual reproduction takes place by simple copulation
of the tips of the multinucleate hyphae. The tips consist of terminal
swellings and arise as branches from the mycelial mats. The tips
are attracted to one another by sex hormones or they merely come into contact
by chance. After contact, each of the hyphal tips swells, and a septal
wall is formed separating the cytoplasm and nuclei in the swollen end from
the rest of the hypha. The wall between the adjacent tips then dissolves,
and there is mixing of the cytoplasm from the two mating strains, followed
by paring of the nuclei. The new cell (zygote), which is the product
of this fusion enlarges and the walls become thick and pigmented.
When nuclear fusion takes place, a diploid nucleus is formed. After
a period of inactivity, the zygote cracks open, a spormgiophore emerges,
and a sporangium develops. The spormgiophores within the sporangium
undergo a nuclear conjugation followed by reduction division to produce
a haploid spore containing genes from both parents.
In
the ascomycetes, sexual reproduction systems are quite varied. However,
when two sexual types (+ and -) come together the end result of all these
variations is the formation of the ascus or sac in which the sexual spores
(ascospores) are produced.
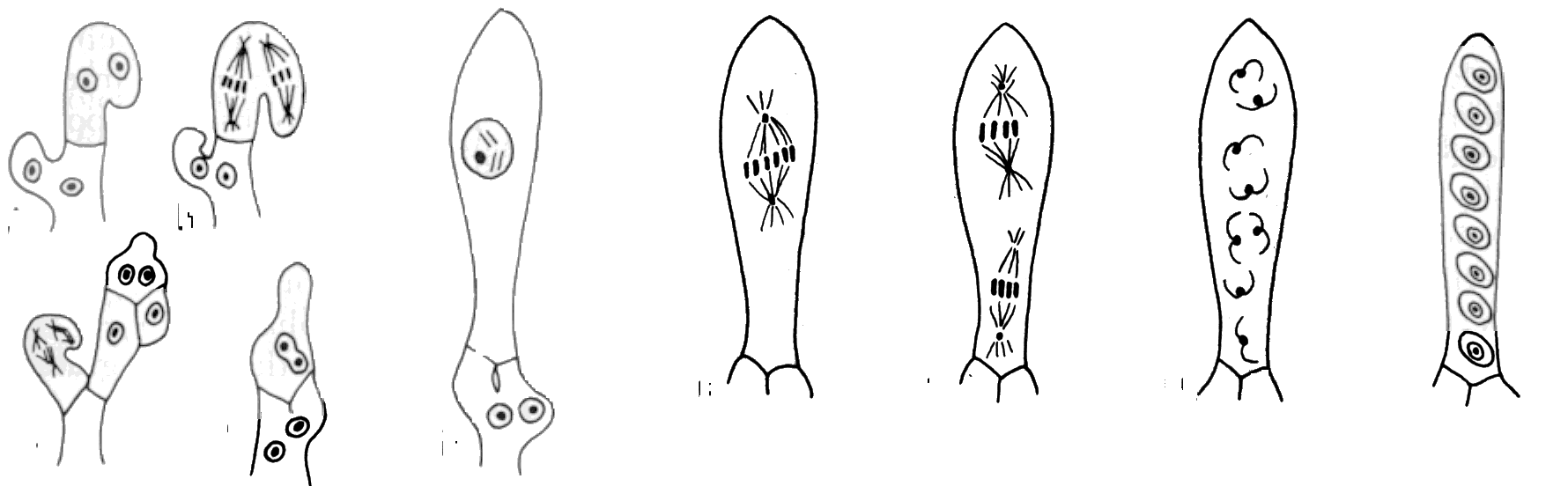
After nuclear conjugation and reduction division have occurred, the ascus contains eight haploid nuclei destined to be contained in spores.
In the basideomycetes sexual reproduction occurs only among those species
which form clamp connections as seen below.
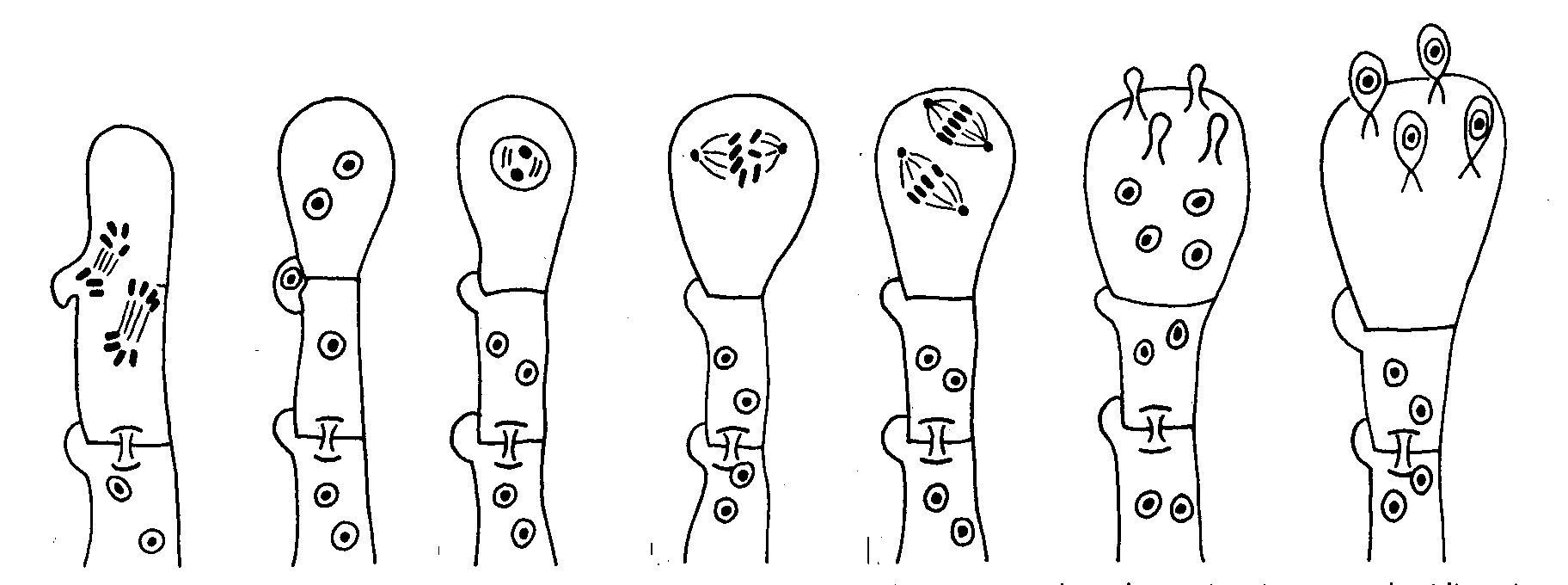
1. Genetic exchange between viruses occurs by recombination, reassortment and polyploidy.
2. Recombination is the breakage and reunion of homologous regions in the nucleic acid of two viruses.
3. Reassortment is an exchange of nucleic acid segments between viruses with segmented chromosomes.
4. Polyploidy is the incorporation of more than one complete genome into the same virus capsid.
5. Genetic exchange between bacteria occurs by transformation, transduction and conjugation.
6. Transformation is the uptake of extracellular DNA by bacteria.
7. Transduction is the transfer of bacterial DNA, by a virus, from one bacterial cell to another.
8. A prophage is a bacterial virus that has the ability to integrate its DNA into the DNA of a bacterial cell.
9. Lysogenization is the process of integrating bacteriophage DNA into bacterial DNA.
10. Lysogenic conversion is a change in the phenotype of a bacterial cell due to lysogenization.
11. In restricted transduction, only those genes near the prophage attachment site are transduced.
12. In generalized transduction, all genes have an equal probability of being transduced.
13. Plasmids are small (relative to the chromosome) autonomously replicating circular pieces of DNA.
14. A plasmid that is capable of integrating into the chromosome is termed an episome.
15. Plasmids may carry genes for transfer of DNA (their own DNA as well as chromosomal DNA) to another bacterial cell, drug resistance, virulence, production of antimicrobial agents and for metabolic activities.
16. Bacterial conjugation is plasmid-mediated gene transfer.
17. A plasmid that can mediate gene transfer is termed the F (fertility) plasmid. A bacterial cell containing the F plasmid is called an F+cell. A bacterial cell not containing a F plasmid is called the F-cell. A bacterial cell containing a F plasmid integrated into the bacterial DNA is termed a Hfr (high frequency of recombination) cell.
18. Bacteriocins are antimicrobics that are active only against other strains of the same species that produced them.
19. In general, plasmids can carry genes coding for drug resistance, virulence, antimicrobial agents, and metabolic activities.
20. Genetic exchange between fungi occurs via exchange of whole nuclei. The method of exchange varies between the Phycomycetes, Ascomycetes and Basideomycetes but the end result is the same, i.e., formation of a zygote.